 吉川泰弘の
ホームページへようこそ!
吉川泰弘の
ホームページへようこそ!
おまけのおまけ:「霜降り肉やトロは何故おいしいか?」
この項目を書いたきっかけは以下に述べました。全部を書き終えてもう一度考えてみると、この表題にいきつきました。
①霜降り肉やトロ(例えば獣肉:牛肉と魚肉、マグロ)がおいしいのは、脳がおいしいと思うからです。ウニ、イクラ、生卵のおいしさも同じです。
②では、何故、脳は霜降り肉やトロや卵をおいしいと思うのでしょうか?
脳は必須アミノ酸(脊椎動物が自分では合成できないで、摂取を必要とするアミノ酸)と
必須脂肪酸(動物性脂肪酸でアラキドン酸とDHAなど)を必要とするからです。
③脳は何故、特に必須アミノ酸と必須脂肪酸を欲しがるのか?
中枢神経系で最も必要な活動は、寝ている間も24時間、中枢神経回路を動かし続けること
で、そのためには糖と酸素を十分供給し、ミトコンドリアでATPを作り続ける必要があ ります。分裂しない神経細胞が使うエネルギーは、神経回路をスクラップ・ビルトするこ
と(可塑性を維持すること、新しい神経回路を作り続けること)です。ヒトの特徴は巨大
な脳を持ったことです。そして、その脳をフルに使ったことです。
④霜降り肉などの特徴は、赤身肉(蛋白質、アミノ酸)と脂肪(必須脂肪酸)を別々に分布させないで、混在させたことです。適切な霜降り状態は、摂取した時、最も理想的な脳への
必須栄養源と言えるのではないでしょうか?
⑤弥生時代以来、農耕民族として生活してきた日本人が、何故霜降り肉のおいしさに目覚め
たのかは、よくわかりませんが、魚(マグロ、ウナギ)やクジラ(海棲哺乳類)の肉と魚
介類の卵の脂肪の生食経験から、牛や豚の品種改良に行きついたのかもしれません。
古くから家畜の飼育経験を持つ欧米人が赤身肉を好むのは、狩猟時代の経験(刷り込
み)が強すぎたのかもしれません。彼らは感染症の経験から生食よりも火を使った加工肉
などの方を選択しました。美味しさよりも安全性を優先する必要があったのでしょう。卵
や魚のような感染病原体の少ない食品を生で食べる文化は、四海を海に囲まれ、温帯で海
産物の多様な日本の環境が影響しているかもしれません。
⑥直接のエネルギー源となる糖(植物由来では澱粉など、動物由来では肝臓のグリコーゲ
ン)、酸素を運ぶ鉄(血液、魚の血合いなど)、そして必須アミノ酸(情報発現のための
核酸合成にも必要)と必須脂質(神経細胞の構成成分)は、ミトコンドリアの活動を通し
て、脳の発達、維持に必要であり、おいしいと感じる素材なのでしょう。また、食べると
いう行為が、細胞が細胞を摂食するととらえれば、魚のような刺身は最高の摂食手段で
す。火を使うメリットは、感染症の防止や食べやすくすることです。しかし、栄養源とし
てみると、高熱により蛋白質等の高分子の変性が起こることを考えると、デメリットもあ
ります。生食文化の日本人(生卵、刺身など)の知恵かもしれません。
ここからが、本論です。「各種動物の必須アミノ酸と食性と脳」
先日、食肉フォーラムの中で、必須アミノ酸と食性と脳の関連について、ちょっとした議論がありました。時間の合間に関連する事項をピックアップしてみました。肉食の重要性は必須アミノ酸の問題よりも脂質にあるのではないかと思われます。
「ヒトは肉好きのサル」と言われています。しかし、動物が、共生体として葉緑体でなくミトコンドリアを選んだ時から肉食を運命づけられました。脳の大半は脂質ですが、構成成分の必須要素は動物由来のアラキドン酸とDHAです。これはミトコンドリアの元祖プロテオ細菌が、TCAサイクルを回すために必要とした栄養素です。プロテオ細菌も葉緑体の元祖であるシアノバクテリアも全アミノ酸を合成できますが、主要な脂質はシアノバクテリアでは植物と同様、リノール酸、リノレン酸などです。これは、後述するようにミドリムシ(ユーグレナ)の生き方で、よくわかります。

1、食性と脳
①食性:一般に肉食性・草食性・雑食性などがある。
肉食性:肉食、魚食、虫食、血食
草食性:草食、花粉食、蜜食、樹液食、果実食、穀物食
雑食性:動物と植物の両方
土食性:土、泥
デトリタス食性:デトリタス
(生物遺体や生物由来の物質の破片や微生物の死骸、あるいはそれらの排泄物を起源とす
る微細な有機物粒子。通常は表面や内部に繁殖した微生物群集を伴う。)
菌食性:真菌、細菌
「食べる」ということを考えてみると、従属栄養(化学合成細菌や光合成細菌のように自らエネルギー生産をして独立栄養で生きられる生物が産生した有機物を横取りして栄養源とする)の原生動物のアメーバやマクロファージ細胞が貪食するイメージがわきます。実際従属栄養化した真核生物単細胞の原生動物は、丸ごと生きた細胞を貪食し、栄養源にしたり(細胞が細胞を食べる)、共生体あるいはゲノムを水平移動させ(宿主のゲノムに取り込む)、進化してきました。
葉緑体をベースとした独立栄養の光合成細菌、藻類、植物系と、ミトコンドリアをベースとした従属栄養の動物系に分岐してきたわけですが、細胞が細胞を食べるという意味では、動物細胞は動物細胞を食べる(肉食性)のが、最も効率が良いということになるでしょう。尤も、ミトコンドリアの直接のエネルギー源となる糖は、植物が合成するので、ビタミン等を含め植物食も必要です。
②ヒトの食性
www.expo-cosmos.or.jp/img/main/kosmos/22/transcripts_22.pdf
「食とは?人類は何を食べてきたか?」 人は肉好きのサルである!
③ヒトはオランウータン、ゴリラ、チンパンジー(類人猿)と共通の祖先から進化しました。約1300万年前にオランウータン、約650万年前にゴリラ、約500万年前にチンパンジーがヒトとの共通祖先から分分岐しました。ヒトの特徴は知能が高いことであり、知能の発達には脳が大きくなることが必須でした。
チンパンジーの脳容積は300ml程度、現代人の成人男性の脳容積の平均は約1350mlです。チンパンジーと同程度の脳容積しかなかった初期人類(猿人:ラミダス猿人)から、高度の知能をもった現生人類に進化する過程で脳容積は4倍以上に増えました。チンパンジーの脳容積は500万年前と同じで、ヒトの脳容積が約4倍も増えた理由は、何故でしょう。ヒトが動物性食糧を多く摂取するようになったからであると考えられます。

ヒトの脳の糖の消費量は、全身の15~20%で、酸素消費量は20%です。脳以外の組織・臓器は残りの80%の消費量で生きています。 チンパンジーの脳のエネルギー消費量は約5%で、残りの95%は、脳以外の臓器・組織が利用します。ヒトの脳は拡大した分、そのままエネルギー消費量を増やしました。エネルギーの獲得競争で、脳は一切妥協しない暴君のようなものです。飢餓状態になった時、各種臓器・組織を犠牲にしても脳は萎縮しません。脳の機能が止められる(一酸化炭素中毒や肺炎、肺水腫など)と、ヒトは容易に死に陥りますが、脳が機能を止めない限り、かなりの飢餓(栄養エネルギー供給不足)でも生存し得ます。
脳組織の50から60%は脂質から構成されています。このうち約3分の1はアラキドン酸やドコサヘキサエン酸のような多価不飽和脂肪酸であり、アラキドン酸は必須脂肪酸で人は体内で合成できません。ドコサヘキサエン酸は同じω3系不飽和脂肪酸のα-リノレン酸から体内で変換されますが、その効率は極めて悪く、最近ではドコサヘキサエン酸も必須脂肪酸に分類されるようになっています。脳の成長に必要なアラキドン酸とドコサヘキサエン酸は食事から摂取しなければならないのですが、これらの脂肪酸は植物性食物には少ししか含まれていません。アラキドン酸は肉、ドコサヘキサエン酸は魚脂に多く含まれています。
アラキドン酸は20:4ω-6、すなわち炭素が20(エイコサ)、シス型2重鎖結合が末端から6番目(オメガ6)から始まり、2重結合が4つ(テトラエン)です。オメガ6・エイコサ・テトラエン酸です。ドコサヘキサエン酸は、同様に22:6ω-3ですから、炭素数が22(ドコサ)、2重鎖結合が末端から3番目から始まり(オメガ3)、6つの2重結合(ヘキサエン)を持つ脂肪酸です。リノール酸は、炭素が18個(オクタデカ)、6番目の炭素から2重鎖が入りますが(オメガ6)、二重鎖は2つ(ジエン)です。リノレン酸は炭素数が18で末端から3番めの炭素から2重鎖が始まり(オメガ3)、2重鎖は3つ(トリエン)です。オメガ3・オクタデカン・トリエン酸ということになります。ちなみに、EPAは「エイコサペンタエン酸」です。イワシやサバ・アジなどの青魚に多く含まれるオメガ3系脂肪酸です。C20:5ω-3になります。ヒトの体内では、ほとんど作ることのできない「必須脂肪酸」です。

オランウータンやゴリラやチンパンジーは今でも熱帯の密林に生息し、植物性食物の多い食事をしています。昆虫や鳥類の卵や小型哺乳類など動物性食物も食べますが、主体は果実、植物の葉、芽、根など糖質の多い食物です。氷河期の間も、アフリカやアジア地域にわずかに残った森林で生き延びました(現在の熱帯雨林は地球のわずか6%弱です)。彼らは、森に残ったためにヒトのような進化をとげられなかったと思われます。ヒトが彼らと違ったのは、どこからでしょうか?ヒトも森林に住んでいたころは植物性の食糧でしたが、ヒトが肉食になったのは、約250万年前から氷河期が始まったためであると思われます。
約440万年前に現在のエチオピア地域の熱帯雨林に生息していた初期人類のラミダス猿人の
食事は、木の葉や果実やベリー類など軟らかい植物性食物が主体でした。歯の構造から硬い植物を食べるようには適応していませんでした。約400万年〜200万年前に生存したアウストラロピテクスは二足歩行を行うようになり、密林からより開けた草原で住むようになりました。アフリカ東部や南部のサバンナの環境に適応し、歯が発達して硬い殻をもつ大きな種子や地下の根なども食べるようになりました。植物性食物を中心にして、さらに小動物の狩猟や、動物の死肉や肉食獣の食べ残しから動物質性食糧を得るようになっていきました。
ヒトが狩猟を開始する直接のきっかけは250万年前くらいから起こってきた気候や環境の変化であると考えられます。このころから、地球は氷河期に移行し、地球上の気温が低下していき、アフリカの熱帯雨林は縮小し、草原やサバンナに変化していきました。氷期の間、地球全体が乾燥し、降雨量が少なくなると大きな樹木は育たなくなり、草原が増えます。草食動物が増え、草食動物を獲物とする大型の肉食動物が棲息するようになります。ヒトはそのような獣を狩猟によって食糧にしてきました。動物以外にも、漁によって魚介類も多く摂取しています。
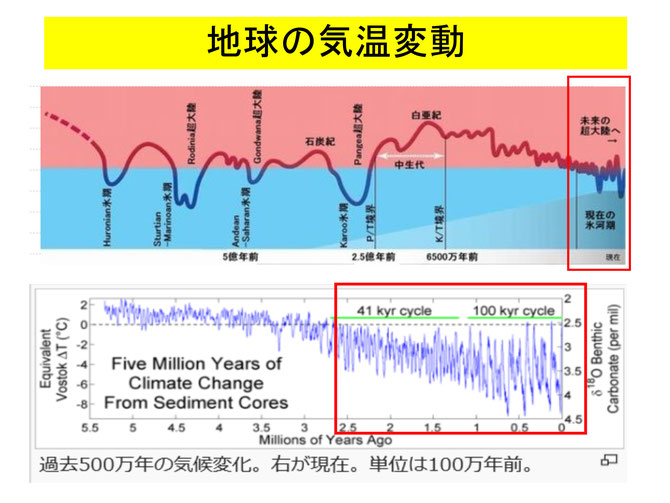
間氷期になって気候が暖かくなって樹木が成長すると木の実や果物なども増えますが、基本的には肉や魚など動物性の食糧が半分以上を占めていました。氷河期は地球の気候が長期にわたって寒冷化する期間で、北アメリカやヨーロッパ大陸に氷床が拡大し、アジアやアフリカも気温が低下して涼しくなり、熱帯雨林は縮小します。氷河期は数万年続いて再び温かい気候に戻ります。氷期と氷期の間を間氷期といいます。
地球は、約250万年以降、4万年から10万年の周期で氷期と間氷期を繰り返しています。最後の氷期が終わったのが約1万年前(農耕の開始)で、現在は間氷期です。ホモ属(Homo:原人)が現れたのは 250万年〜200万年前です。この頃から人類は石器を道具として利用し、狩猟や肉食獣の食べ残しから得た動物性の食糧が増えてきました(この時期に脳が大きくなったようです、600~800ml)。さらに、160万年前くらいから人類は火を使うようになり、食物を火で加熱することによって栄養の吸収が良くなりました。150万年前に住んでいたホモ・エレクトスは積極的に狩猟を行っていたようです(このころのジャワ原人の脳容積は800~1200mlです)。 このように初期人類の食事は、植物性食料由来の糖質が多いものでしたが、250万年くらい前から動物性食料が増えるようになり、少なくとも150万年前くらいから農耕が始まる1万年前くらいまでは、低糖質・高蛋白食でした。
約1万年前に最後の氷河期が終わり地球は温暖化し農耕と牧畜が始まりました。農耕によって穀物の摂取が増えました。糖質の摂取量は現代人では1日250から400g程度ですが、狩猟採集時代の糖質摂取量は1日10から125gと推定されています。農耕が始まってから、成人の平均身長は減少しているという報告があります。農耕が始まってから、人類の歴史の中ではじめて脳の重量が減少していることが報告されました。
現代人の脳容積は、2万数千年前までヨーロッパに存在したネアンデルタール人(旧人:脳容積は1200~1700ml)、その後新人である、クロマニョン人20万年前に出現し、その脳容積は、1400~1500mlとなりました。現生人類(6万年前にアフリカ大陸から拡散)の脳容積は、彼らより10%程度小さいことが明らかになっています。その理由としてタンパク質や不飽和脂肪酸の摂取量の減少が指摘されています。農耕によって穀物が豊富になり、糖質が増えた分、肉や脂肪の摂取量が減ったからであると考えられています。
www.daiwa-pharm.com/info/fukuda/7096/(一部改変)

2、必須アミノ酸と食性、進化
①必須アミノ酸の必須性は、炭素骨格によります。必須アミノ酸の合成は、非必須アミノ酸の合成経路に比べて、長いステップが必要です。進化の過程で、この合成系を失ったと考えられます。原核生物、真菌類、植物はすべてのアミノ酸合成能を持っています。 後述するように、例外はありますが、ヒトを含むすべての動物と原生生物(真核単細胞)の細胞性粘菌は必須アミノ酸を持っています。
②必須アミノ酸の種類はほとんど同じです。原生生物の進化のある時に、ほぼ10種のアミノ酸合成能を一斉に失いました(ゲノムからも明らかで、炭素骨格の合成酵素を欠損した)。しかし、どの過程で能力を失ったか、その原因は何か?は不明です。他方、進化の分岐からすると、多細胞生物の祖先(襟鞭毛虫)は、全アミノ酸合成能を持っています。2胚葉性の刺胞動物(サンゴ)も、全アミノ酸を合成します(後述)。他方、3胚葉の土壌線虫は必須アミノ酸を持っています(甲殻類、節足動物の祖先)。進化のいろいろな時点で、別々に必須アミノ酸(合成系の欠損)が生じたとも考えられます?
③原生動物の進化は、トリコゾア亜門(ジアルジア)、キネトプラスト門(トリパノソーマ、ユーグレナ)、アメーバ界(アメーバ)、アピコンプレックス門(胞子虫)、繊毛虫門(繊毛虫)の順です。

④肺炎の原因となるレジオネラ.ニューモフィラ(L.pneumophila)は、アミノ酸のような単純な栄養素が不足している水生環境において、複雑なバイオフィルム内または自由生活のアメーバ細胞内で生存する環境微生物です。 L.ニューモフィラは、細胞内成長に必要なアミノ酸を絶対に必要とするにもかかわらず、Arg、Cys、Ile、Leu、Met、Thr、Valに対して栄養要求性です(Pineら、1979; Georgeら、1980 ; Reevesら、1981; Ristrophら、1981; TeshおよびMiller、1981; Teshら、1983; MolofskyおよびSwanson、2004; Wielandら、2005; Eylertら、2010)。 驚くべきことに、水棲環境におけるL.ニューモフィラの主要な宿主アメーバであるAcanthamoebaeは, Arg, Ile, Leu, Val (IngallsおよびBrent、1983)に対し栄養要求性であり、アミノ酸栄養要求のL.ニューモフィラと重なっています。また、別のアメーバ宿主であるDictyostelium discoideumは、L.ニューモフィラのアミノ酸栄養要求性と重複する11種類のアミノ酸に対して、やはり栄養要求性です(Payne and Loomis、2006)。 D. discoideumはシステインは栄養要求性ではありませんが、このアミノ酸が培養時に培地に存在しない場合、その増殖率および生産率は有意に低下します(FrankeおよびKessin、1977)。 L.ニューモフィラによるアミノ酸栄養要求性とその自然宿主であるアメーバとの同期は、偶然である可能性は低いと思われます。従属栄養細菌L.ニューモフィラは、もともと必須アミノ酸要求性であったか?or アメーバと共生する間に宿主同様に遺伝子を欠損させたか?は不明です。Environ Microbiol. 2014 Feb; 16(2): 350–358
⑤アブラムシとブフネラ菌の栄養共生:アブラムシの必要な必須アミノ酸は10 種類です(アルギニン,イソロイシン,スレオニン,トリプトファン,バリン,ヒスチジン,フェニルアラニン,メチオニン,リジン,ロイシン)。アブラムシが常食とする植物の師管液は,ショ糖は多く含みますが,アミノ酸の組成は片寄っており,アブラムシの必要とする栄養素を満たしていません。ブフネラ菌は,アブラムシが師管液から得たり、代謝老廃物として過剰にあるアミノ酸(グルタミンやアスパラギン)をもらい、アブラムシが合成できない必須アミノ酸を提供します。リケッチア,クラミジア,マイコプラズマなどの細胞寄生細菌は,アミノ酸合成に関わる遺伝子をほとんど失い,アミノ酸は宿主から横取りしています。 ところが,ブフネラ菌は多くの点で宿主に依存していながら,アミノ酸合成については大腸菌の持つ遺伝子の約半分(アブラムシの必須アミノ酸を合成する遺伝子)を残しています。残っているのは,アブラムシが合成できないアミノ酸をつくる遺伝子ばかりです。ブフネラ菌の祖先は大腸菌と同じようにすべてのアミノ酸を合成できたのですが,共生してから,アブラムシの合成できるものはそちらに依存することになったのだろうと思われます。今ではブフネラ菌と宿主であるアブラムシが,お互いに相手の合成できないアミノ酸を供給し合う,持ちつ持たれつの関係が成立しているといえます。
⑥ユーグレナ(ミドリムシ)は、非常に興味深い生物です。植物・動物の両性質を持ちますが、その脂質代謝では、独立栄養条件(光合成可)で生育するとリノール酸、リノレン酸が多く、従属栄養条件(暗条件)ではアラキドン酸やエイコサペンタエン酸が多く含まれます。リノール酸、リノレン酸は葉緑体に、アラキドン酸、エイコサペンタエン酸はミトコンドリアに分布します。また、葉緑体欠損株では、光照射に関係なくアラキドン酸等が生成されます。光合成可、暗条件、光合成欠損株でもアミノ酸合成には差がない(必須アミノ酸はなく、全アミノ酸合成能を持っています)。

⑦ジアルジア(鞭毛虫)では、エネルギー代謝の副生成物としてのアラニンの合成とは別に、デノボ合成が報告されている唯一の他のアミノ酸はバリンです。 したがって、ジアルジアはほとんどのアミノ酸の合成能に欠けており、栄養型(トロフォゾイト)では、それが増殖する腸内環境から必須アミノ酸を掻き集めることに依存しています。(寄生性として退行性進化をした。)
Clin Microbiol Rev. 2001 Jul; 14(3): 447–475.7
⑧多細胞生物の起源(立襟鞭毛虫は必須アミノ酸合成可能):必須アミノ酸の合成に必要な経路は動物で失われた?様です。 これらの経路には、ロイシン、バリン、イソロイシン、ヒスチジン、リシン、スレオニン、トリプトファン、フェニルアラニンおよびメチオニンが含まれます。アミノ酸合成は、動物共通祖先より襟鞭毛虫においてより完全です。 紫:襟鞭毛虫(Urchoanozoan)と動物共通祖先(Urmetazoan)の両方に存在するアミノ酸経路。 青:アミノ酸経路が襟鞭毛虫に存在し、動物共通祖先に欠けている(図1)。系統樹分岐とは一致しません!より古いトリコゾア亜門の鞭毛虫(寄生性の原虫、ジアルジアは退行性進化した結果アミノ酸合成能を欠損?)。自由生活(非寄生性)の従属栄養真核生物としてキネトプラスト門の鞭毛虫(ユーグレナ)及び襟鞭毛虫(多細胞生物の祖先)は必須アミノ酸合成能を持っています。
https://www.biorxiv.org/content/biorxiv/early/2017/12/26/211789.full.pdf
⑨多細胞動物でのアミノ酸合成能。動物は代謝ニーズを満たすのに合成できない、あるいは十分な量合成できないアミノ酸を食事に頼っている。これらは所謂「必須アミノ酸」である。この必須アミノ酸のセットは、脊椎動物と多くの無脊椎動物の間で似ている。これまで、最も原始的な無脊椎動物である刺胞動物のアミノ酸合成能についての情報はなかった。本研究では、イシサンゴ目の代表的な刺胞動物によるアミノ酸合成能を調べた。 サンゴ礁サンゴ、星の砂サンゴ、シカツノサンゴ、ハマサンゴなどを対象とした。トリプトファンまたはシステインを検出することができず、グルタミン酸とグルタミン、アスパラギン酸とアスパラギンを区別することもできなかった。しかし、通常、動物に必須であると考えられている8つのアミノ酸は、テストされた5種類のサンゴで作られていた(少量でしか作られなかったものもあった)。これらは、バリン、イソロイシン、ロイシン、チロシン、フェニルアラニン、ヒスチジン、メチオニン、リシンである。これらのアミノ酸を合成する刺胞動物の能力は、他の後生動物(メタゾア、metazoan)から刺胞動物が分岐した進化の歴史の指標となる可能性がある。

⑩3胚葉、線虫のアミノ酸合成能:土壌線虫は自由生活にもかかわらず、必須アミノ酸が存在し、ほぼ脊椎動物と同じである。線虫では数種類のビタミン摂取が必要である。チアミン、リボフラビン、葉酸、ナイアシンアミド、パントテン酸、ピリドキシン(Dougherty et al。、1959; Nicholas et al。、1962; Dougherty and Hansen、1957; Lu et al。、1974) 、ビオチン(Nicholas and Jantunen、1963)およびビタミンB12(Luら、1976)である。 アミノ酸では、アルギニン、ヒスチジン、リシン、トリプトファン、フェニルアラニン、メチオニン、スレオニン、ロイシン、イソロイシン、バリンは、 土壌線虫C. elegansやC. briggsae の増殖を維持するために、必須アミノ酸である(Vanfleteren、1980)。 (Nicholas et al。、1960; Rothstein and Tomlinson、1961; 1962; Rothstein、1963; 1965)。 対照的に、アラニン、アスパラギン酸、システイン、グルタミン酸、グルタミン、グリシン、セリン、およびチロシンは必須アミノ酸ではない(Vanfleteren、1980)。www.wormbook.org/chapters/www.../intermetabolism.html
⑪直接の祖先である魚類:アミノ酸のうち10種類は魚類では合成できない必須アミノ酸である。 食餌によって供給されなければならない10種類は、メチオニン、アルギニン、スレオニン、トリプトファン、ヒスチジン、イソロイシン、リジン、ロイシン、バリン、フェニルアラニンである。 これらのうち、リジンとメチオニンは多くの場合、最初の制限アミノ酸である。 植物性(大豆ミール)蛋白質で調製された魚の飼料は、メチオニン含量が低く、最適な成長と健康を促進するためには、大豆ミールをベースとした飼料には、メチオニンを追加する必要がある。
⑫ウシの必須アミノ酸はアルギニン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、スレオニン、トリプトファン、バリンの10種類です。ただし、胃の微生物が合成するので、少ない量でいい?あるいはいらないという意見もある。ブタでは9種の他アルギニンも必須アミノ酸。鶏ではアルギニン、グリシンも必須アミノ酸である。


進化と必須アミノ酸の存在(アミノ酸合成能)
|
エネルギー代謝 |
独立栄養 |
従属栄養(宿主) |
寄生生物 |
|
|
化学合成 光合成 |
|
|
|
細菌 |
硝酸菌 光合成細菌
全合成 |
大腸菌
全合成 |
マイコプラズマ クラミジア 無合成 クレブシエラ 必須アミノ酸 |
|
原生生物 |
藻類 鞭毛虫:ユーグレナ 全合成 |
鞭毛虫:ユーグレナ 葉緑体欠損:全合成 立襟鞭毛虫 多細胞起源:全合成 根足類:アメーバ 必須アミノ酸 繊毛虫 必須アミノ酸 |
鞭毛虫 ジアルジア ほぼ無合成
|
|
真菌類 |
|
全合成 |
|
|
動物:2胚葉 |
|
刺胞動物:サンゴ 全合成 |
|
|
3胚葉 |
|
土壌線虫:線形動物 (C. elegance) 必須アミノ酸 |
|
|
節足動物 |
|
アブラムシ 必須アミノ酸 |
共生:ブフネラ菌 必須アミノ酸 のみ合成 |
|
棘皮動物
|
|
ヒトデ、ウミユリ、 必須アミノ酸 |
|
|
脊索動物 |
|
ホヤ、ナメクジウオ 不明 必須アミノ酸? |
|
|
脊椎動物 |
|
魚類、両生類、 爬虫類、鳥類、 哺乳類 必須アミノ酸 |
|
おまけ:
①必須アミノ酸9~10種は、メチオニン、スレオニン、トリプトファン、リジン、ヒスチジン、イソロイシン、ロイシン、バリン、フェニルアラニン、アルギニン、
②植物性蛋白質のアミノ酸組成と動物性蛋白質のアミノ酸組成には大きな差がある。必須アミノ酸の中でリジン、メチオニン、スレオニン、トリプトファンなどが、動物性蛋白質に比べ相対的に少ない。
③分岐鎖アミノ酸(branched-chain amino acids、BCAA)は、分枝(任意の炭素原子に2以上の別の 炭素原子が結合)のある脂肪族側鎖を有するアミノ酸である。タンパク質を構成するアミノ酸では、ロイシン、イソロイシン、バリンの3種の分枝鎖アミノ酸がある。 必須アミノ酸であり、筋タンパク質中の必須アミノ酸の35%を占め、哺乳類にとって必要とされるアミノ酸の40%を占める。分枝鎖アミノ酸はアシルCoA誘導体に変換され、これは続いてアセチルCoAもしくはスクシニルCoAとなり、最終的にクエン酸回路に組み込まれる。
④生命誕生に必要な蛋白質合成には10種ほどのアミノ酸で十分(?)だった。グルタミン酸(アスパ ラギン酸、リジン、アルギニン)、グルタミン(アスパラギン)、プロリン、アラニン、スレオニン(セリン、グリシン)、メチオニン(システイン)、トリプトファン、フェニルアラニン(チロシン)、分岐鎖アミノ 酸群(ロイシン、イソロイシン、バリン)、ヒスチジン?
妻と作った人形。
娘の修学旅行の写真をもとにしました。
妻と作った人形。
娘の修学旅行の写真をもとにしました。

オリジナルの写真です

娘のドイツ時代のカーニバルの写真です。大家さんは子ネズミちゃん「モイスヒェン」といっていました。
下の人形は妻の作品です。


先日、妻の作品が創刊700号記念家庭画報大賞の佳作に入りました。
題「何して遊ぼう」です。


妻が、稽古に通い、粘土で作った作品です。昨年、東京フォーラムで、他の生徒さんと一緒に展示されました、「仙人草」
(水やり不要です)。

妻の人形作品です。
ドイツ時代の香代の幼稚園の友達です

ある夏のスナップです。妻の父母、娘、甥たちの集合写真から作りました。
